
This research is driven by personal criteria - by who is related to me or to my wife - so it is hard to summarise exactly what is here. The detail also varies. Some families I cover in enormous detail. Other families I merely sketch since they are covered by others elsewhere. (Some lines here connect to landed, titled and Royal families.)
I started a website in 1994, but I think I did not start putting genealogy onto it until 1995. There are surviving archive.org copies of my site from 1999. (There are no earlier archive.org copies because the servers it was on before this blocked spiders.)
I think my site is the oldest continuously-running genealogy site in Ireland. (Paddy Waldron's site is older, but took a break.)
A nice way to get an idea what is on the site is to Google image search "site:humphrysfamilytree.com". Here are four rather random surveys of what is on the site:

This all began in 1983, as research in motoring history, when I began to research a series of cars built by my grandfather and others in Ireland in the 1920s and 1930s: The "Thomond" motor car. This led on to researching My family tree in general.
There was a vast amount of family history material to start with because so many of my family were involved in the Irish revolution of 1916-23. Family members were arrested, jailed, on hunger-strike, active in underground government, shot, and even killed. It is a cliché in Ireland that one's ancestors were in the GPO in the Rising in 1916, but my grandfather Dick Humphreys did fight in the GPO in 1916, and his uncle "The O'Rahilly", one of the leaders of the rebellion, was the only leader to be killed in action (the others were executed).
I could summarise my family as almost archetypal Catholics made good in the 19th century in Ireland - after the ending of the repressive laws against Catholics, when a vast body of Irish Catholics slowly began to work their way up from nothing through trade, farming and business, to form the middle class of the emerging state at the start of the 20th century. The O'Maras, O'Rahillys and Flanagans all rose like this over the course of the 19th century. Indeed, the writer Kate O'Brien used the O'Maras as the archetypal example of such a family. My great-great-grandfather Stephen O'Mara was a Parnellite MP. My great-grandfather James O'Mara, Home Rule MP, Sinn Fein TD, was a member of the 1st Dail in 1919, as was my grand-uncle W.T. Cosgrave, who became the first Prime Minister of an independent Ireland in 1922. His son Liam Cosgrave (my mother's first cousin) became Prime Minister of Ireland in 1973.

I never planned to research My wife's family tree as well, but it was too interesting to resist, and again, a lot of source material has survived which I could read.
Her family descends from an impossibly romantic story: Augustus "Reebkomp", the illegitimate son of Henry Herbert, 10th Earl of Pembroke in England in the 18th century. Reebkomp was the product of an infamous elopement in 1762 that is referred to in the movie The Madness of King George. He grew up with his legitimate half-brother (the future 11th Earl), and money came out of the Earl's family for him and his descendants for almost a full hundred years. Through him, my wife descends from most of the great old Anglo-Irish families of the Pale of the last few hundred years, which is of particular interest to me.
Apart from the above, her family history can be summarised as merchants and traders in Scotland in the 17th and 18th centuries, who rose to the middle class there centuries before that was possible in Ireland. Indeed, through one line we see a slow descent from a great family, Skene of Skene, Aberdeenshire (as opposed to the sudden and spectacular descent of the illegitimate son above).
This is not that unusual. My most popular web page is my collection of Royal Descents of famous people, which shows how much of the population of the West descends provably from medieval European royalty.
See Proof of our descent from Blennerhassett for the proof that we descend from the Blennerhassett Baronets branch.
I call it as proved that my ancestor George Cashel (born 1807) descends from Robert Blennerhassett (died 1765) and his wife Frances Yielding. Nothing else explains the DNA. That we descend from this family is proven, but the exact descent is unproven. It remains to prove this from paper, to confirm from paper records what the DNA says must exist.
This means that I too connect to the World family tree. I too have a Royal Descent. The most recent monarch I descend from is Edward III.
Much of this earlier part of my site, such as my sketches of medieval royalty and other well-known families, is not original research, nor intended to be, but is merely a kind of extended Ancestors Chart for my wife and I.
This new theory is explained at my popular Common ancestors of all humans page.
And permanent copies of the files will be at the
Internet Archive.
This entire site (including images and video) is being archived there:

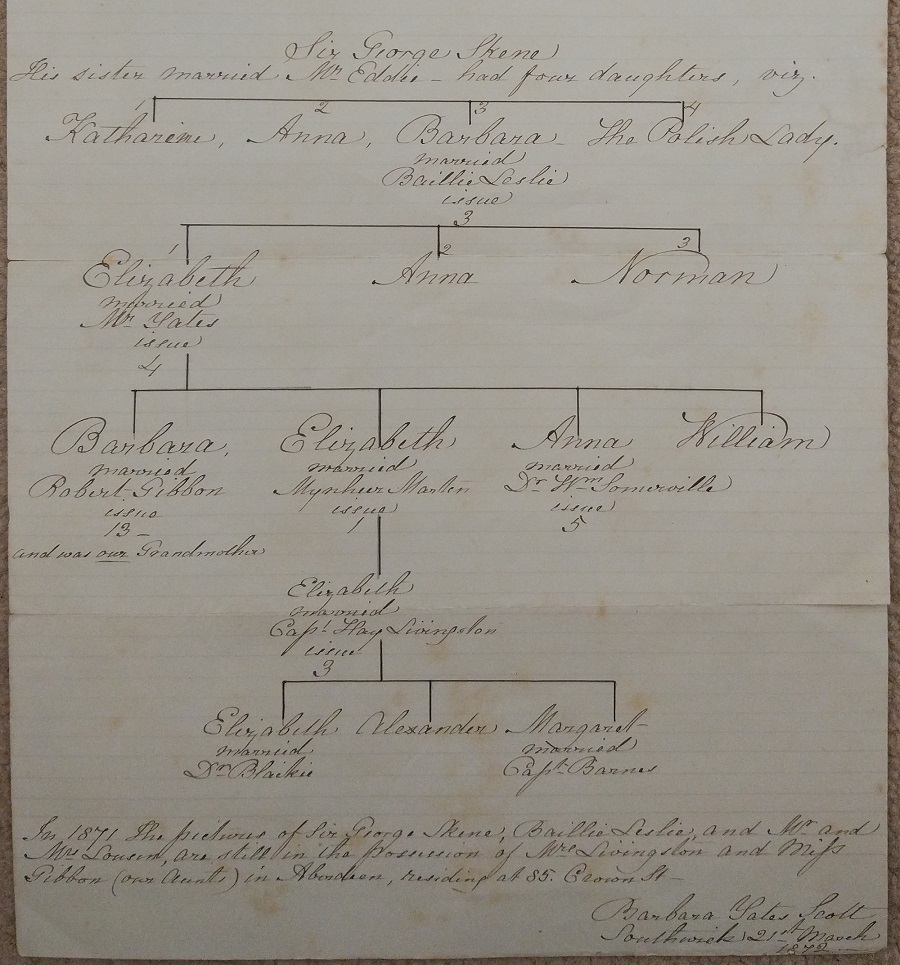
Sometimes things are easier.
From this
single sheet found in an old tin box
could be reconstructed and corroborated
a descent of one of my wife's ancestors
from the great Scottish family of Skene,
leading ultimately to a Royal Descent.
"I spent the whole evening reading over a parcel of my own letters, addressed to my sisters
which they fancied worth preserving.
What a strange creature is man, more strange still woman!
I am as different to what I was 15 years ago,
as I am now to an entire stranger.
My letters must have been amusing at the time they were written
but the subjects were so personal that now when time has shown me those persons in a different
point of view, and changed the whole face of things,
they appeared to me highly ridiculous,
and my own compositions not worth reading again.
I threw the whole bundle into the fire"
- Diary of Ann Gibbon, Sun 23rd Jan 1820.
"One happy day in my life"
- Charles Morris, 1891.
"there's a blacktop road;
a faded yellow centerline;
It can take you back to the place;
But it can't take you back in time."
- The song
Flies on the Butter (You Can't Go Home Again) (2004).
See video.
Please donate to support this site.
I have spent a great deal of time and money on this research.
Research involves travel and many expenses.
Some research "things to do"
are not done for years, because I do not have the money to do them.
Please Donate Here
to support the ongoing research and
to keep this website free.
